











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo

 Permalink
Permalink
1. Introduction
Efficiency indicators in milk systems are affected by several factors which could be related to individual characteristics. Interindividual differences in animals’ behavioral responses to different environmental circumstances are defined as temperament 1)(2) 3. For dairy cows, temperament has been associated with productivity (milk yield, quality, and milk ability) 4)(5)6)(7) ; metabolic features 8)(9) , and behaviors such as adaptation to a novel milking system 10)(11) . However, the knowledge about what determines or modulates temperament remains limited. Genetics is a factor that influences the cows’ temperament, since differences between dairy breeds have been reported 12)(13) 14. Additionally, the differences in productivity among breeds could be, in part, explained by temperament differences 12)(15) 16. In Uruguay, North American (NA) Holstein is the most common dairy strain (78%), but cows of New Zealand (NZ) origin (13%) have increased17. Previous studies have reported that NZ cows produced lower individual milk yield but higher milk component concentrations18, spending more time eating total mixed ration19 and having greater energy and feed efficiency than NA cows20, although both had a similar economic-productive breeding index. A possible explanation for the differences in milk yield between cows of different temperament is the human-animal relationship 21)(22) . Therefore, differences in milk yield between NZ and NA could be attributed to variations in their reactivity to humans, such as milking reactivity, across both strains. The genetic differences in temperament between strains could be due to history of pasture breeding and selection based on productivity and body size of each one 23)(24) . Additionally, the feeding system could be another factor that affects the cow’s reactivity. In pasture-based management systems, human-cow interaction is typically lower than in indoor housing systems, which could result in cows under grazing systems showing greater reactivity in tests that assess fear of humans, such as increased flight distance 25)(26) 27. According to our knowledge, studies comparing the cow’s temperament of the different breeds or by different strains within the same breed are scarce. Thus, we hypothesize that the NZ cows would have greater reactivity to different temperament tests than NA ones, and cows under pasture system will show greater reactivity than cows under pasture+partial mixed ration. The objective of the present study was to compare the responses to different temperament tests of NA and NZ Holstein cows maintained in two different feeding strategies (pasture or pasture+partial mixed ration).
2. Materials and methods
An experiment study18 was carried out with 120 Holstein cows over a one-lactation period at the experimental station of the National Agricultural and Livestock Research Institute (in Spanish, INIA), Colonia, Uruguay (34°20′ S, 57°41′ W). All procedures were approved by the Ethics Committee of INIA (file number INIA.2019.11). A full description of the experimental design was included in Stirling and others18. Four groups were derived from the combination of two feeding strategies: Grass Maximum or Grass Fixed, and two Holstein strains: NZ or NA. The Grass Maximum and Grass Fixed feeding strategies differed in the proportions of grazed pasture in the diet with the same level of concentrate per cow. The cows in Grass Fixed were fed an allowance of ⅓ pasture, ⅓ concentrate, and ⅓ silage. The silage was offered as a partial mixed ration on a concrete and dirt feedpad (4.5 m2/cow), and the total area (feeding and rest area) was 240 m2 per cow. The cows in Grass Maximum had a flexible pasture allowance determined by the pasture growth rate, which was estimated weekly, offering whole-crop silage on a feedpad as a buffer in case of pasture shortage. The pasture was a mix of Festuca arundinacea, Dactylis glomerata, Medicago sativa, and Lolium multiflorum. The commercial concentrate was offered daily and individually in the milking parlor.
Thirty NA and 30 NZ cows were randomly assigned to each feeding strategy before calving, ensuring that groups were balanced for expected calving date, parity, milk yield, and milk fat content from the previous lactation. Finally, of the 60 cows in each Holstein strain cow temperament was assessed in a total of 48 NZ cows (primiparous: 12, multiparous: 36) and 50 NA cows (primiparous: 19, multiparous: 31), in the milking parlor (herringbone type) using a milking reactivity score (MR), and in the handling pen using flight speed and flight distance tests (details given below). The tests in the milking and handling pen were performed on the same day at 107.5 ± 30.3 and 173.8 ± 32.1 days in milk (two repetitions). First, the MR was assessed during afternoon milking, and then the cows were transferred to the handling pen where the other tests were performed, by the same observer. Milking reactivity was measured using a method modified from Sutherland and Huddart28. Each cow was scored based on the stepping and kicking behaviors using an 8-point scale during preparation for milking and attachment of the milking cluster: (1) no hind foot movement, (2) one or two slow and gentle (hoof elevated at less than 15 cm above the ground) hind limb movements, (3) three or more slow and gentle hind limb movements in inconstant rhythm (not moving during the whole procedure), (4) three or more slow and gentle hind limb constant movements (during the whole procedure), (5) vigorous hind limb movements (elevating hooves 15 cm above the ground), (6) vigorous and constant hind limb movements, (7) kicking the milker, elevating the hind hoof above hock line and directing it laterally, and (8) the cow presented high reactivity and had to have one or both hind limbs tied to enable the procedure to be performed. The observation was made by one observer positioned behind the animal in the milker pit, being able to observe both hind legs.
Flight speed (FS) was measured using a method modified from Gibbons and others29. The time taken by each cow to exit the squeeze chute and cover a distance of 2.7 m was recorded using a purpose-built device that measured exit speed. The unit comprised two light beams and reflectors and one readout unit. Once the cow passed through both light beams, the exit time was recorded in seconds and converted into speed, in m/s. The handler stood approximately 1 m away from the side of the crush and behind the shoulder of the animal during the test. Flight distance (FD) was measured using a method modified by Waiblinger and others26. The distance (in m) that cows allowed a non-familiar person to approach before expressing the first withdrawal response was recorded. The test was performed with each cow individually kept in a pen of 131 m2. After the cows exited the squeeze chute, the observer remained stationary on the opposite side of the pen entrance, approximately 8 m away from the cow, waiting for the cow to stand still before starting the test. Then, the person approached the animal slowly (one step per s), with her hands down and arms held close to the body. When the cow expressed any withdrawal reaction (i.e., the animal moved both forelimbs), the observer stopped and measured the distance to the cow's nearest front hoof using an odometer (MW40M, Stanley, USA).
Cows were weighed individually and fortnightly after the morning milking throughout lactation using a portable electronic scale (Farmquip Ltd., Hawke's Bay, New Zealand). All statistical analyses were carried out with SAS software (version 9.4, SAS Institute Inc., Cary, NC, USA). Univariate analyses were performed to identify outliers and to verify the normality of residuals, and homogeneity of variances was verified with Levene’s test. Considering that the day of assessment (107.5 and 173.8 days in milk) did not affect the cow’s reactivity, we averaged the two repetitions by each test to obtain a single value per cow for the analyses. General linear models (GLM procedure) were used for the dependent variables FS and FD, and a generalized linear model with gamma distribution and inverse link function (GENMOD procedure) was used for MR. Fixed effects of Holstein strains (NZ and NA), parity (primiparous and multiparous), feeding system (Grass Maximum and Grass Fixed), and all two-factor interactions were included. Non-significant interactions (α>0.05) were excluded from the final models. In addition, body weight average was compared between primiparous and multiparous cows by GLM procedure. Post-hoc analysis was performed using Tukey´s test. The α values were considered significant when ≤0.05, and the data are presented as least square means ± SEM.
3. Results and discussion
For MR, the main factors of strain and parity were not significant, but their interaction affected cows’ reactivity (P=0.05). The difference on MR between NZ and NA was observed only for multiparous cows, with higher milking reactivity for NZ than NA (Table 1). The Holstein strains differed for FD (P=0.03), with longer flight distances for NZ (4.2 ± 1.3 m) than for NA (3.7 ± 1.1 m) cows, without parity effect. The FS was not affected by both factors studied (1.18 ± 0.3 m/s; mean ± SD). Finally, the feeding system did not affect the cow’s reactivity nor their interactions with strain.
Table 1: Milking reactivity (MR) of primiparous and multiparous Holstein cows of two genetic strains (North American - NA, and New Zealand - NZ)
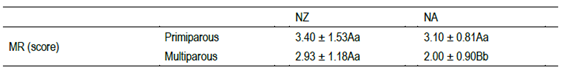
Different letters in the same column are main statistical differences between Holstein strain in the same cow parity (capital letters, p≤0.05), and between parity in the same strain (lowercase letters p≤0.05).
To our knowledge, this is the first study evaluating the relationship between temperament tests and Holstein genetic strains. The current study showed that NZ cows were more reactive to humans in both milking (MR) and paddock (FD) than NA cows. However, at milking this difference was only shown for multiparous cows. Differences for temperament among dairy breeds have been reported in previous studies30. For example, in a study conducted in New Zealand Jersey, cows were regarded as more “placid” during milking than Holstein cows of NZ origin31. In its turn, Mpisana and others32 compared crossbred Jersey × Friesian with purebred Friesian cows in South Africa, showing that the crossbreds had greater avoidance distance but lower milking reactivity than the Friesian cows reared in the same environment. To our knowledge, this is the first study to examine temperament differences of NZ and NA breeding lines. As evidenced in our study, cows of different genetic lines cannot only differ in physical and productive parameters 18)(20)23)(24) , but also in their behavior. The genetic selection could explain the greater flight distance and milking reactivity in NZ than in NA cows, since both lines were breeding under different environmental circumstances and with different goals. The effects of selection primarily for some traits (e.g. productivity or body size) could be related to the temperament genomic loci 30)(33) 34. On one hand, for Holstein cows of NA origin the genetic correlation between milking temperament and milk flow was reported, as well as genetic correlations between bull EBV for milking temperament and milk flow and with various reproduction, conformation, and auxiliary traits35. On the other hand, the NZ cows have been selected for smaller body size23, which could be associated with reactive temperament34. Additionally, cows of lower weight or smaller sizes could be characterized as more “nervous/anxious”32, since smaller animals would have more room to move in the milking parlor36. This also might be an explanation for our findings, since our NZ cows were smaller than NA18. The NZ cows were also more reactive than NA cows for MR test, but this was observed only in multiparous animals. First, we could speculate that for primiparous, the novel milking environment would lead to high milking reactivity, making it difficult to observe differences between strains (NZ or NA). In fact, previous studies reported that primiparous Holstein cows had higher incidence of stepping and kicking during milking than multiparous ones 4)(6) 37. It is interesting to highlight that for NZ cows we did not observe MR differences between primiparous and multiparous.
Based on the results, we suggest that NA cows reduce their reactivity over time, which would not be the case of NZ cows. The strain differences in MR could also be attributed to variations in body size, which only differed between multiparous cows of both strains. In primiparous, the body weight did not differ for both strains (NZ= 450.3 ± 13.5 Kg, NA= 505.6 ± 51.5 Kg, P= 0.09). However, in multiparous, NZ cows were lighter (544.0 ± 58.5 Kg) than NA multiparous cows (625.4 ± 68.1 Kg) (P<0.0001), explaining the strains differences only for multiparous cows. Finally, it is interesting to note the result that the feeding system did not affect the temperament of the cows. In this sense, not only the different interaction with humans, but also the characteristics of the feed itself in composition as well as in other behavioral parameters (such as grazing) did not greatly influence temperament in our study.
Beyond our results, future studies would be important to evaluate whether different feeding systems can affect temperament by considering other variables, such as animal density, feeding management characteristics (for example, 100% confinement and 100% pasture systems), as well as different diets, among other factors. The advantages of a herd with calm animals could be fewer human accidents and less stressed cows during milking, therefore, fewer negative effects on milk production. However, this could be true in conventional milking systems, as automatic milking systems (AMS) are recommended for reactive animals11. In this sense, future research should determine the reactivity and productivity of both Holstein strains in other milking systems, such as the AMS. In addition, reducing the susceptibility of dairy cows to be frightened should improve their ability to adapt to various farm environments and, consequently, enhance farm economy and animal welfare.
4. Conclusions
In conclusion, dairy cows’ genetic strain and parity were factors related to temperament in adult Holstein cows, mainly in MR and FD tests. In the milking parlor, multiparous NZ cows were more reactive than NA cows, what did not happen for the primiparous ones, for which the variations between both strains were not evident. The NZ cows were more reactive to humans than NA cows in the paddock, exhibiting longer flight distances.

