










Servicios Personalizados
Revista

Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Links relacionados
Compartir

 Permalink
PermalinkAgrociencia (Uruguay)
versión impresa ISSN 1510-0839versión On-line ISSN 2301-1548
Agrociencia Uruguay vol.20 no.2 Montevideo dic. 2016
Caracterización del comportamiento reproductivo de dos nuevos híbridos de mandarina
Mautone Ana Paula1, Rey Florencia1, Pereira das Neves Virginia1, Guimaraes Natalia1, Gambetta Giuliana1, Gravina Alfredo1
1Universidad de la República, Facultad de Agronomía, Departamento de Producción Vegetal. Avenida Garzón 780, 12900 Montevideo, Uruguay. Correo electrónico: agravina@fagro.edu.uy
Recibido: 21/12/15 Aceptado: 10/11/16
Resumen
Durante dos ciclos anuales se estudió el comportamiento reproductivo de dos nuevos híbridos de mandarinas (tangor ‘Ellendale’ x Satsuma ‘Owari’), denominados M9 y B30, desarrollados en el Programa de Mejoramiento Genético de Citrus de Uruguay. Ambos demostraron presentar autoincompatibilidad, tanto en autopolinización natural como en polinizaciones dirigidas. El polen de ambos híbridos presentó baja capacidad de germinación in vitro y en el caso de M9, a partir del tercer día post-antesis, se evidenció pérdida de viabilidad in vivo de sus óvulos. En condiciones de libre polinización, M9 desarrolló un 80 % de frutos aspermos, mientras que en B30, el porcentaje alcanzó apenas a 16 %. Polinizado con polen de mandarina ‘Afourer’, naranja ‘Valencia’ y limón tipo ‘Lisbon’, M9 solamente con ‘Afourer’ desarrolló frutos con semillas (33%). B30 por el contrario, presentó 100 % de frutos con semillas al ser polinizada con ‘Afourer’ y ‘Valencia’. Se discuten las diferencias reproductivas entre ambos híbridos.
Palabras clave: AUTOINCOMPATIBILIDAD, CAPACIDAD PARTENOCÁRPICA, SEMILLAS, VIABILIDAD DE POLEN
Summary
Characterization of the Reproductive Behavior of Two New Mandarin Hybrids
The reproductive behavior of M9 and B30, two new hybrids of mandarin (‘Ellendale’ tangor x Satsuma ‘Owari’) developed at the Uruguayan Citrus Breeding Program, was studied during two annual cycles. Self-incompatibility of both cultivars was demonstrated, both in natural self-pollination and in directed pollination. In both hybrids, low in vitro pollen germination ability was found. In M9, since 3rd day after anthesis, an in vivo ovule viability loss was evident. At open pollination, 80 % of M9 fruit was seedless, while in B30 the percentage scarcely reached 16 %. After pollinated with ‘Afourer’ mandarin, ‘Valencia’ orange and ‘Lisbon’ lemon, M9 had only 33 % of seedy fruit when ‘Afourer’ was the pollinator. On the contrary, B30 presented 100 % of seedy fruit after being pollinated with ‘Afourer’ and ‘Valencia’. Reproductive differences between both hybrids are discussed.
Keywords: SELF-INCOMPATIBILITY, PARTHENOCARPIC ABILITY, SEEDS, POLLEN VIABILITY
Introducción
La tendencia actual en el desarrollo de variedades de Citrus indica que una de las principales características requeridas en la fruta para consumo en fresco, es la ausencia o bajo número de semillas. Aunque ese atributo no es estricto o absoluto y ha variado en el tiempo y en los diferentes mercados, una variedad puede considerarse sin semillas si el número de estas no excede a una cada doce frutos. Con ese objetivo, se han desarrollado en los últimos años programas de mejoramiento genético tendientes a lograr cultivares de alta calidad organoléptica que desde el punto de vista reproductivo sean autoincompatibles, y/o que no polinicen ni sean polinizados por otros genotipos mediante la polinización cruzada (Vardi et al., 2008; Handaji et al., 2010; Navarro et al., 2015).
La autoincompatibilidad en Citrus -incapacidad de producir la fecundación en condiciones de autopolinización- es fundamentalmente de tipo gametofítico (Soost, 1969; Distefano et al.; 2009). Los ejemplos más conocidos de cultivares partenocárpicos y autoincompatibles son las naranjas de ombligo, las mandarinas Satsumas y las Clementinas (Frost y Soost, 1968; Soler, 1999). En nuestras condiciones, Inzaurralde et al. (2010) demuestran la autoincompatibilidad del tangor ‘Ortanique’, el cual es eficientemente polinizado por la mandarina ‘Nova’ (Borges et al., 2009). También la mandarina ‘Afourer’ se comporta en Uruguay como autoincompatible, pero es polinizada por varios cultivares, por lo que sus frutos presentan semillas en condiciones de polinización abierta (Gravina et al., 2011; Gambetta et al., 2013).
El polen de los cítricos es pesado y con capacidad de adherencia, por lo que la polinización entre cultivares compatibles es fundamentalmente entomófila, siendo las abejas los insectos de mayor importancia en el proceso. Aunque se ha demostrado que el polen puede ser transportado varios cientos de metros por las abejas (Chao et al., 2005), su eficiencia en la polinización y posterior presencia de semillas se va perdiendo, disminuyendo de un rango de 52- 80 % a 13-39 % de frutos con semillas entre los primeros 60 m y los 100-250 m de distancia de la fuente de polen (Otero y Rivas, 2010). En el caso de cultivares autoincompatibles pero no estériles, las alternativas para obtener frutos sin semillas son principalmente el aislamiento, ya sea geográfico o mediante mallas antiabejas colocadas sobre las plantas durante la floración (Nadori, 2006).
En el marco del Programa de Mejoramiento Genético de Citrus en Uruguay (Rivas et al., 2014), se han obtenido varios híbridos de mandarina, entre los cuales se destacan por la calidad de su fruta los denominados M9 y B30 (‘Ellendale’ x Satsuma ‘Owari’). Para la zona de Salto, M9 alcanza su maduración en julio con buen tamaño de fruta y 11,5 °Brix; B30 madura en agosto superando los 15 °Brix con tamaños similares a M9. Observaciones preliminares indican que en condiciones de libre polinización, ambos presentan semillas, aunque en diferentes proporciones. En este trabajo se planteó como objetivo la caracterización del comportamiento reproductivo de M9 y B30 con énfasis en su tipo de compatibilidad, capacidad partenocárpica y formación de semillas en condiciones de polinización libre y dirigida.
Materiales y métodos
Los experimentos se realizaron en la Estación Experimental San Antonio, de la Facultad de Agronomía, ubicada en el departamento de Salto, Uruguay (31° 21’ LS, 57° 45’ LO), en los ciclos productivos 2013-2014 y 2014-2015.
En los tres experimentos se utilizaron plantas adultas de M9 y B30 (tangor ‘Ellendale’ x Satsuma ‘Owari’), injertadas sobre Poncirus trifoliata L. Raf. y citrange Q VII (C. sinensis L. Osb. x P. trifoliata), respectivamente. Los árboles se encontraban en una colección varietal del programa de mejoramiento genético de la Facultad de Agronomía. Los árboles de mandarina ‘Afourer’, naranja ‘Valencia’ y limón tipo ‘Lisbon’, utilizados en el Experimento 3 como donadores de polen, se encontraban en el mismo predio.
Experimento 1. Estudio de la autocompatibilidad y aptitud partenocárpica
En plantas de M9 y B30 se marcaron diez ramas secundarias de similar tamaño por cultivar, con un promedio de 217 y 161 flores respectivamente. A cinco ramas, se les colocaron estructuras de alambre, las cuales se cubrieron con malla antiabejas durante todo el período de floración. Finalizado el mismo, las mallas fueron retiradas y quincenalmente se evaluó el número de frutitos presentes hasta el final de la caída fisiológica. Con esos datos, se construyó la curva de abscisión de flores y frutitos en cada situación y cultivar y se determinó el porcentaje de cuajado final. En la madurez se contabilizó el número de semillas por fruto.
Experimento 2. Germinación de polen in vitro
Se colectaron 20 flores al inicio de antesis y se colocaron en sílica gel durante 24 h para promover la apertura de las anteras. Posteriormente se hidrataron los granos de polen en heladera a 4 ºC. La siembra se realizó en cámara de flujo laminar sobre portaobjetos conteniendo el medio de cultivo Brewbaker y Kwack (1963) solidificado con Phytagel al 1 % y cubierto con 30 µL de medio líquido. Los preparados se colocaron en cámara oscura a 25 ºC y 80 % de humedad durante 72 h y posteriormente se fijaron con solución de FAA (5 % formol aldehído, 5 % ácido acético, 90 % etanol al 70 %). La germinación de polen se evaluó contabilizando siete repeticiones de 300 y 400 granos de polen por placa para M9 y B30, respectivamente, utilizando un microscopio óptico (Nikon E100). Se consideró grano de polen germinado cuando el tubo polínico duplicaba el tamaño del grano.
Germinación de polen, crecimiento del tubo polínico y viabilidad de óvulos in vivo
En cada cultivar se marcaron y embolsaron 90 brotes de flor terminal en estado 59 de la escala BBCH (Agustí et al., 1995). A los 0, 3, 6, 9 y 12 días se colectaron 15 flores por cultivar y se fijaron en solución FAA, conservándose a 4 ºC hasta su observación en microscopio de fluorescencia (Olympus Vanox AH3 y filtro U BH2-DMU para el rango de emisión Hg 334-365 nm). Los pistilos fueron seccionados en ovario y estilo-estigma, y colocados en una solución de sulfito sódico (5 %) durante 12 h. Luego de este período se enjuagaron en agua destilada y se colocaron en una nueva solución de sulfito sódico. El ablandamiento de los tejidos se realizó en microondas (15-20 segundos, 700 W). Las secciones de los estilos se tiñeron con azul de anilina al 0,1 % en K2HPO4 0,1 N (Kearns e Inouye, 1993), y se colocaron en portaobjetos con una gota de glicerina para evitar la deshidratación. La evaluación de la germinación de los granos de polen en el estigma se realizó con microscopio de fluorescencia, contabilizando los granos germinados por flor en un total de 200-270 granos por fecha y cultivar. Para evaluar la viabilidad de los óvulos, los ovarios reblandecidos se seccionaron ecuatorialmente y bajo lupa binocular se rescataron los óvulos con la ayuda de pinzas. Estos fueron colocados en portaobjetos y para teñirlos, se les agregó unas gotas de solución de azul de anilina al 0,1 % en tampón K2HPO4 0,1N (Kearns e Inouye, 1993). Luego de 20 minutos, se secaron con papel de filtro, se les agregó glicerina, se cubrieron con cubreobjetos y se realizó un ‘squash’. Los preparados fueron observados en microscopio de fluorescencia. La viabilidad de óvulos se determinó contabilizando al azar, en promedio 70 y 38 óvulos por fecha en M9 y B30 respectivamente, considerándose no viables aquellos que presentaron depósito de calosa en la zona de la chalaza, emitiendo fluorescencia.
Experimento 3. Estudio de polinización cruzada
Previo a la antesis, se seleccionaron para cada tratamiento 80 y 100 brotes de flor terminal en M9 y B30, respectivamente. Al alcanzar el estado 59 de la escala BBCH, se aplicó con un pincel polen de naranja ‘Valencia’, limón tipo ‘Lisbon’, mandarina ‘Afourer’ y polen propio, obtenido en forma similar a lo descripto en el Experimento 2 y posteriormente se cubrieron con bolsas de tul. A los 15 días se retiraron las bolsas y quincenalmente se cuantificó el número de ovarios/frutitos presentes, hasta el final de la caída fisiológica, para determinar el cuajado final. Además, para estudiar la viabilidad de los granos de polen de los genotipos M9 y B30, se colectaron 10 flores en el estado 59, y la extracción del polen se realizó según lo descrito anteriormente. Con un pincel fino se espolvoreó el polen sobre un portaobjeto que contenía una gota de solución al 0,5 % de cloruro de tetrazolio y 10 % de sacarosa; se cubrió inmediatamente con portaobjeto para excluir el oxígeno, y se incubó a 60 ºC durante 1 h (Bolat y Pirlak, 1999). En microscopio óptico se evaluó la viabilidad del polen, considerando los granos teñidos de rojo como viables, y los amarillos como no viables.
Resultados
Nuestros resultados indican que en condiciones de autopolinización, el 98 % y el 100 % de los frutos de los híbridos M9 y B30, respectivamente, no presentaron semillas, por lo que puede afirmarse que ambos genotipos son autoincompatibles. Por el contrario, en condiciones de polinización abierta, el comportamiento reproductivo fue diferente, ya que en M9, el 80 % de los frutos no presentó semillas, mientras que en B30 ese porcentaje alcanzó solamente el 16 % (Cuadro 1). Adicionalmente, el número promedio de semillas por fruto en condiciones de polinización abierta fue muy bajo en M9 (0,57 semillas por fruto) y de los frutos con semillas un 73 % presentaron solo una o dos semillas. En las mismas condiciones, B30 presentó 6,4 semillas por fruto y solamente un 16 % de estos tuvo una o dos semillas. Esto indica que M9, aun en condiciones de libre polinización, presenta alta capacidad para cuajar frutos aspermos, lo que constituye una característica de alto valor comercial.
La dinámica de abscisión de flores y frutitos fue diferente entre ambos híbridos y condiciones de polinización. Al retirar las mallas 40 días post-antesis, M9 presentaba un mayor cuajado que B30, aunque durante el período de abscisión las diferencias disminuyeron, finalizando en valores similares (Figura 1). En M9, durante todo el período evaluado, el porcentaje de cuajado fue levemente superior en las ramas en condiciones de autopolinización, en relación al testigo de polinización abierta. Por el contrario, en B30 el porcentaje de cuajado con o sin malla se mantuvo en niveles similares hasta mediados de diciembre, pero durante la fase final de abscisión el cuajado en polinización abierta fue levemente superior a la autopolinización (Figura 1). El cuajado final en ambos cultivares fue relativamente elevado, sin diferencias significativas entre autopolinización o polinización abierta, aunque con tendencias diferentes. En M9, el cuajado final bajo malla fue levemente superior al de polinización abierta, mientras que en B30 ocurrió lo contrario (Cuadro 1).
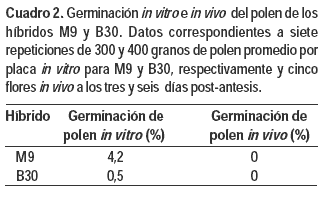
Ambos cultivares presentaron baja capacidad de germinación de polen in vitro, en especial B30, que fue inferior a 1 %. La germinación in vivo fue nula en ambos híbridos hasta los 12 días post antesis (Cuadro 2).
La viabilidad de los óvulos también fue diferente en ambos híbridos. En M9 a partir del día 3 post-antesis (DPA) se evidenció deposición de calosa en la chalaza, la cual se incrementó hasta los 12 DPA. En B30 los primeros síntomas de pérdida de viabilidad se observaron a los nueve días post-antesis (Cuadro 3).
Los resultados de los tratamientos de polinizaciones dirigidas, en términos de porcentaje de frutos con semilla y número de semillas por fruto se presentan en el Cuadro 4. En M9 solamente se obtuvieron frutos con semilla cuando sus flores fueron polinizadas con polen de mandarina ‘Afourer’. Esto confirma tanto la autoincompatibilidad de este híbrido como la escasa capacidad de sus flores de ser fecundadas con polen foráneo. Por el contrario, los frutos de B30 presentaron semillas con todos los polinizadores utilizados, alcanzando el 100 % con mandarina ‘Afourer’ y naranja ‘Valencia’ (Cuadro 4). La viabilidad de los granos de polen de naranja ‘Valencia’ alcanzó el 10,5 %, el de ‘Afourer’ 8,5 %, el de limón tipo ‘Lisbon’ el 18,2 %, el de M9 13,5 % y el de B30 19 %.
La aptitud partenocárpica de ambos híbridos fue diferente cuando sus flores fueron emasculadas, o emasculadas y polinizadas manualmente con su propio polen. El cuajado fue nulo en M9 y B30 en flores emasculadas, pero con autopolinización M9 alcanzó 6,7 % mientras que B30 solamente cuajó un 2,5 % de sus flores (Cuadro 5). La respuesta a los cultivares polinizadores utilizados también fue diferente entre los híbridos y se relacionó con la presencia de semillas. Con polen de ‘Afourer’ y ‘Valencia’, M9 presentó un porcentaje de cuajado relativamente bajo, inferior inclusive al tratamiento de autopolinización, mientras que B30 cuajó cinco y cuatro veces más, respectivamente (Cuadro 5).
Discusión
La autoincompatibilidad en cítricos es una característica de alto valor comercial, ya que en cultivos aislados de polinización cruzada permite la obtención de frutos sin semillas. Varios cultivares de mandarinas e híbridos (Clementinas, Satsumas, ‘Nova’, ‘Ortanique’, ‘Afourer’) presentan esta característica y por lo tanto la capacidad de producir frutos partenocárpicos (Soost, 1956; Hearn et al., 1969; Soler, 1999; Gambetta et al., 2013). De acuerdo a los resultados obtenidos, proponemos que los híbridos M9 y B30 son autoincompatibles, ya que no presentaron semillas ni en condiciones de autopolinización, ni cuando fueron polinizados manualmente con su propio polen.
En condiciones de polinización abierta, en una parcela con un elevado número de variedades y por lo tanto una fuerte presión de polen, M9 presentó solamente 20 % de frutos con semillas y una media de 0,6 semillas por fruto. Un estudio similar en nuestro país determina que la mandarina ‘Tango’, una mutación generada a partir de la mandarina ‘Afourer’ patentada en California, presenta el 12 % de los frutos con semillas y una media de 0,15 semillas por fruto en condiciones de polinización abierta (Gravina, sin publicar), asemejándose a lo encontrado en M9. Por el contrario B30, en condiciones de polinización abierta, solamente alcanzó un 16 % de sus frutos sin semillas.
La escasa germinación de polen in vitro y nula in vivo de ambos híbridos, parecen no relacionarse con factores de clima prevalentes antes y durante la antesis, ya que en los 10 días previos a la obtención del polen para su análisis, la temperatura media fue 20,2 °C y la HR 82 %, que pueden considerarse adecuadas para la germinación del polen de acuerdo a Distefano et al. (2012). Temperaturas menores a 10 °C durante los días previos a la antesis han sido reportadas como críticas para la germinación del polen in vitro (Pardo et al., 2007, 2010), lo que no ocurrió durante el período considerado. A su vez, esa baja germinación no permitió determinar el tipo de autoincompatibilidad que presentan ambos híbridos. Hasta el momento, en el género Citrus solamente ha sido reportada la autoincompatibilidad de tipo gametofítico (Soost, 1969; Distefano et al., 2009; Gambetta et al., 2013), caracterizada por la germinación de los granos de polen en el estigma y el desarrollo parcial del tubo polínico en el estilo, sin sobrepasar el 50 % de la longitud del mismo.
En M9 la viabilidad de los óvulos comenzó a disminuir desde el tercer DPA, incrementándose la deposición de calosa a los 12 DPA, lo que puede explicar la baja capacidad de formación de semillas en condiciones de libre polinización. Este comportamiento sugiere un período efectivo de polinización de corta duración, lo que limitaría su capacidad de formación de semillas, similar al reportado por Mesejo et al. (2007) en mandarina Satsuma ‘Owari’. En B30, recién a los 9 DPA se observaron óvulos con signos de pérdida de viabilidad, lo que puede explicar al menos parcialmente la alta capacidad de formar frutos con semillas de este híbrido en condiciones de libre polinización.
Nuestros resultados indican que M9 presenta partenocarpia estimulativa, ya que las flores emasculadas presentaron una abscisión total, no pudiendo cosecharse frutos, mientras que las polinizadas manualmente con su propio polen, alcanzaron un 6,7 % de cuajado, desarrollando todos sus frutos sin semillas. A su vez, en ramas aisladas, el porcentaje de cuajado fue similar al obtenido en condiciones de polinización abierta. La capacidad partenocárpica de B30, resultó menor a la de M9, tanto en condiciones de aislamiento de polinización cruzada, como polinizada manualmente con su propio polen. De acuerdo a su habilidad de cuajar frutos en ausencia de polinización cruzada, los cultivares autoincompatibles se pueden clasificar en improductivos o de baja capacidad partenocárpica, y productivos o de elevada capacidad de cuajado (Guardiola, 1992). Las causas de la falla en el cuajado inicial son fundamentalmente el bajo contenido de giberelinas en los ovarios (Talón et al., 1990, 1992; Mesejo et al., 2013) y la menor capacidad de atraer los carbohidratos en condiciones de alta competencia (Gómez- Cadenas et al., 2000; Iglesias et al., 2003).
El estudio del comportamiento con polinizaciones dirigidas demostró que M9 presenta una baja capacidad de formación de semillas con polen de ‘Afourer’ y frutos aspermos con polen de naranja ‘Valencia’. B30 por el contrario, presentó el total de sus frutos con semillas en las polinizaciones con mandarina ‘Afourer’ y naranja ‘Valencia’, mientras que con limón tipo ‘Lisbon’ alcanzó el 67 %. Considerando las temperaturas medias ocurridas entre 10 días pre-polinización y 10 días post-polinización, las mismas se ubicaron en 16,2 °C el experimento en M9 y 14,7 °C en el B30, por lo que la baja presencia de semillas en el primero no debe atribuirse a factores exógenos. Las polinizaciones dirigidas con polen compatible promueven en general un elevado cuajado de frutos (Borges et al., 2009; Gravina et al., 2011), y aún más, considerando que se seleccionan brotes de flor terminal, que naturalmente presentan mayor capacidad de cuajado (Erner y Shommer, 1996; daCunha Barros y Gravina, 2006; Rivas et al., 2007). Esta fue la situación en B30, especialmente con polen de ‘Afourer’ y ‘Valencia’. Inesperadamente, en M9 el porcentaje de cuajado con los mismos polinizadores fue bajo e inferior al alcanzado en condiciones de autopolinización, lo que sugiere la existencia de algún mecanismo de esterilidad parcial en este híbrido. Esto confirma los resultados del primer año donde en condiciones de autopolinización las flores de M9 presentaron igual o más capacidad de cuajado que en condiciones de polinización abierta.
Considerando en su conjunto las características reproductivas de los híbridos en estudio, ambos presentan autoincompatibilidad. M9 se presenta como muy promisorio para la producción de frutos con pocas semillas o aspermos, aun en condiciones de producción comercial en convivencia con otras variedades. B30, por el contrario, en condiciones de polinización abierta presenta muy buena intercompatibilidad y un alto número de semillas, por lo que de implantarse comercialmente deberán tomarse las precauciones para su aislamiento de cultivares que sean fuente de polen compatible.
Agradecimientos
Este trabajo se realizó en el marco del proyecto FMV_2_2011_6997, financiado por la Agencia Nacional de Investigación e Innovación (ANII).
Bigbliografía
Agustí M, Zaragoza S, Bleiholder H, Buhr L, Hack H, Klose R, Staub R. 1995. Escala BBCH para la descripción de los estadios fenológicos del desarrollo de los agrios (Género Citrus). Levante Agrícola, 332: 189 – 199.
Bolat I, Pirlak L. 1999. An investigation on pollen viability, germination and tube growth in some stone fruits. Turkish Journal of Agriculture and Forestry, 23(4): 383 – 388.
Borges A, daCunha Barros M, Pardo E, García M, Franco J, Gravina A. 2009. Cuajado de frutos en tangor Ortanique en respuesta a la polinización y a distintas situaciones de estrés. Agrociencia (Uruguay), 13(1): 7 – 18.
Brewbaker JL, Kwack BH. 1963. The essential role of calcium ion in the pollen tube growth. American Journal of Botany, 50(9): 859 – 865.
Chao C-CT, Fang J, Devanand PS. 2005. Long distance pollen flow in mandarin orchards determined by AFLP markers, implications for seedless mandarin production. Journal of the American Society for Horticultural Science, 130(3): 374 - 380.
daCunha Barros, Gravina A. 2006. Influencia del tipo de brote en el cuajado y crecimiento de frutos del tangor ‘Ortanique’. Agrociencia (Uruguay), 10(1) : 37 - 46.
Distefano G, Hedhly A, Las Casas G, La Malfa S, Herrero M, Gentile A. 2012. Male–female interaction and temperature variation affect pollen performance in Citrus. Scientia Horticulturae, 140: 1 – 7.
Distefano G, Caruso M, La Malfa S, Gentile A, Tribulato E. 2009. Histological and molecular analysis of pollen-pistil interaction in clementine. Plant Cell Report, 28: 1439 - 1451.
Erner Y, Shomer I. 1996. Morphology and anatomy of stems and pedicels of spring flush shoots associated with citrus fruit set. Annals of Botany, 77: 537 – 545.
Frost HB, Soost RS. 1968. Seed reproduction : development of gametes and embryos. En: Reuther W, Batchelor LD, Webber HJ [Eds. ]. The Citrus Industry. Vol. 2 Berkeley : University of California Press. pp. 290 - 319.
Gambetta G, Gravina A, Fasiolo C, Fornero C, Galiger S, Inzaurralde C, Rey F. 2013 Self-incompatibility, parthenocarpy and reduction of seed presence in ‘Afourer’ mandarin. Scientia Horticulturae, 164: 183 – 188.
Gómez-Cadenas A, Mehouachi J, Tadeo FR, Primo-Millo E, Talón M. 2000. Hormonal regulation of fruitlet abscission induced by carbohydrate shortage in citrus. Planta, 210: 636 – 643.
Gravina A, Fornero C, Galiger S, Inzaurralde C, Fasiolo C, Gambetta G. 2011. Partenocarpia, polinización cruzada y presencia de semillas en mandarina Afourer. Agrociencia (Uruguay), 15(2): 40 – 47.
Guardiola JL. 1992. Fruit set and growth. En: 2nd International Seminar on Citrus Physiology; 10 - 13 agosto, 1992; Bebedouro, San Pablo, Brasil. Bebedouro : Editora Legis Summa. pp. 1 – 30.
Handaji N, Arsalane N, Ben Azouz A, Benyahia H, Gaboneet F, Srairi I, Esagide A. 2010. A diversification program of mandarin and orange varieties in Morocco. En: XI International Citrus Congress; 26 - 30 octubre, 2008; Wuhan, China. Beijin : China Agricultural Press. pp. 59 – 61.
Hearn CJ, Reece PC, Fenton R. 1969. Self-incompatibility and the effects of different polen sourcesupon fruit characteristics of four Citrus hybrids. En: Chapman HD. [Ed.]. Proceedings of the 1st International Citrus Symposium. Riverside : University of California. pp. 183 – 187.
Iglesias D, Ibáñez R, Tadeo F, Primo-Millo E, Talón M. 2003. La disponibilidad de carbohidratos mejora el cuajado de los frutos de los cítricos. Levante Agrícola, 365: 160 – 166.
Inzaurralde C, Fasiolo C, Fornero C, Galiger S, Chouza X, Gambetta G, Gravina A. 2010. Autoincompatibilidad, capacidad partenocárpica y mejora del cuajado en tangor ‘Ortanique’ [Cd- Rom]. En: VI Congreso Argentino de Citricultura; 2 - 4 junio; Tucumán, Argentina. pp. 37 – 39.
Kearns CA, Inouye DW. 1993. Techniques for pollination biologists. Niwot : University of Colorado Press. 538p.
Mesejo C, Yuste R, Martínez-Fuentes A, Reig C, Iglesias DJ, Primo-Millo E, Agustí M. 2013. Self-pollination and parthenocarpic ability in developing ovaries of self-incompatible Clementine mandarins (Citrus Clementina). Physiologia Plantarum, 148: 87 – 96.
Mesejo C, Martínez-Fuentes A, Reig C, Agustí M. 2007. The effective pollination period in ‘Clemenules’ mandarin, ‘Owari’ Satsuma mandarin and ‘Valencia’ sweet orange. Plant Science, 173: 223 – 230.
Nadori E. 2006. Nadorcott mandarin : a promising new variety. En: X International Citrus Congress; 15 - 20 febrero, 2004; Agadir, Marruecos. Agadir: International Society of Citricultura. pp. 356 – 359.
Navarro L, Aleza P, Cuenca J, Juárez J, Pina J, Ortega C, Navarro A, Ortega V. 2015. The mandarin triploid breeding program in Spain. Acta Horticulturae, 1065: 389 - 395.
Otero A, Rivas F. 2010. Producción de semillas y métodos de control en el tangor ‘Afourer’ en el litoral norte de Uruguay [Cd-Rom]. En: III Simposio Investigación y desarrollo tecnológico en Citrus; 15 - 17 noviembre, 2010; Salto, Uruguay. pp. 96 – 99.
Pardo J, Bermejo A, Cano A, Zaragoza S. 2010. La temperatura, la viabilidad del polen y la formación de las semillas en los cítricos. Levante Agrícola, 399(1): 20 – 29.
Pardo J, Bermejo A, Cano A, Zaragoza S. 2007. La germinación de polen y la formación de las semillas en los cítricos. Levante Agrícola, 384(1): 16 - 20.
Rivas F, Laxague J, Menes R, Suarez D, Varela P, Spina M, Luque E, Pintos P, Vignale B. 2014. Nuevas oportunidades varietales para la Citricultura : Mandarinas M9, M19 y B30 [Cd-Rom]. En: I Congreso Latinoamericano de Investigación y Desarrollo Tecnológico en Citrus; 3 - 5 noviembre, Salto, Uruguay. pp. 1 – 4.
Rivas F, Gravina A, Agustí M. 2007. Girdling effects on fruit set and quantum yield efficiency of PSII in two Citrus cultivars. Tree Physiology, 27: 527 – 535.
Soler J. 1999. Reconocimiento de variedades de cítricos en campo. Valencia : Generalitat Valenciana. 188p. (Serie divulgación Técnica ; N° 43).
Soost RK. 1969. The incompatibility gene system in Citrus. En: Chapman HD. [Ed.]. Proceedings of the 1st International Citrus Symposium. Riverside : University of California. pp. 189 – 190.
Soost RK. 1956. Unfruitfulness in the Clementine mandarin. Proceedings of the American Society for Horticulturae Science, 67: 171 – 175.
Talón M, Zacarías L, Primo-Millo E. 1992. Gibberellins and parthenocarpy ability in developing ovaries of seedless mandarins. Plant Physiology, 95: 1575 – 1581.
Talón M, Zacarías L, Primo-Millo E. 1990. Hormonal changes associated with fruit set and development in mandarins differing in their parthenocarpic ability. Physiologia Plantarum, 79: 400 – 406.
Vardi A, Levin I, Carmi N. 2008. Induction of seedlessness in Citrus : from classical techniques to emerging biotechnological approaches. Journal of the American Society for Horticultural Science, 133(1): 117 - 126.

