











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Curriculum ScienTI
Curriculum ScienTI
 Permalink
PermalinkIntroduction
Soybean (Glycine max L. Merrill) is one of the major Brazilian agricultural crops, with a production of more than 86 million metric tons of grain in the 2013/2014 season, grown in an area of over 30 million hectares (Conab, 2014).
Among the factors that explain the success of this crop in the country, seed quality ranks high, since it represents the vehicle through which the plant breeder’s creations are incorporated into new cultivars and made available to farmers (Peske, Barros & Schuch, 2012). The amount of seed obtained from the plant breeding process is never enough to meet the market demand, so that its multiplication for several generations is always necessary prior to marketing. Thus, the multiplication process involves branding the successive seed generations derived from the Genetic seed as Basic, Certified 1, Certified 2, S1 and S2, each one as a result of multiplying the previous category, as established by the Law 10.711 of 2003 (Presidência da República, 2003).
Within this scheme of seed production, the responsibility for each stage relies on different players of the supply chain, so that the plant breeder company remains responsible for the production and multiplication of the Genetic seed, while the Basic seed category multiplication is the responsibility of both, the plant breeding company and the seed producer. The latter must be registered at RENASEM, and is responsible for the multiplication of the Certified seed, classes C1 and C2 (Matsuo et al., 2013).
Those responsible for seed multiplication can employ two strategies to achieve their goals, focusing either on the cropping area necessary to produce the estimated tonnage or by manipulating the multiplication factor (MF). While the approach on the cropping area seeks to obtain the maximum possible seed yield per unit of land in one growing season, the MF focuses on obtaining the maximum possible number of seeds per plant. Both approaches to seed production are the result of soybean plasticity, i.e. its ability to adapt to different environmental conditions and crop management practices such as row spacing and plant density, which modify plant morphology as well as yield components (Pires et al., 2000; Rambo et al., 2003).
According to Schuch and Peske (2008), the intraspecific competition generated by the proximity between plants is quite high in a high productivity crop, so that in low populations the yield per plant is higher whereas in high populations the yield of grains per plant is lower. Peixoto et al. (2000) reported that the number of pods per plant is a trait that adjusts to plant density variations, responding in an inverse relationship to the number of plants per unit area. Ludwig et al. (2011) found that the increase in soybean plant density per unit area decreased the number of pods per plant and the number of seeds per pod, but increased the 1000-grain weight, with the highest yields occurring at the highest number of pods per m².
Knowing to what extent seeding density can be reduced to produce individual plants carrying more seeds, i.e. increase the multiplication factor, is extremely useful to breeding programs and seed producers responsible for the multiplication of higher seed categories. This is due to the limited amount of genetic and basic seeds. The search for alternatives for a single seed to multiply and generate the largest possible number of progeny is a major achievement. The objective of this work was to compare the effects of low soybean seeding rates on the multiplication factor (MF), the yield components, and the physiological quality of the seed produced.
Materials and Methods
Seeds of soybean cultivar BMX Potência RR were seeded in plots with eleven rows, spaced at 0.5 m and eleven meters in length. The effective area for each plot comprised 9 m², where the outer row on each side of the plot, as well as a length of 0.5 m from each border, were discarded. To target final plant densities of 35, 50, 65, 80, 95 and 110 thousand plants per hectare, eighteen seeds per meter of the row were seeded, and thinning was performed after plant emergence. A completely randomized block design with four replicates was laid on a field adjacent to a commercial crop in the outskirts of the city of Capão do Leão, state of Rio Grande do Sul, Brazil.
Plant protection treatments were performed whenever necessary, using a knapsack sprayer with 20 liters of spray volume, following the same procedure for all experimental units.
Sample plants were manually collected when seeds reached 18 % moisture content and artificially dried to 12 % moisture content, proceeding to manual threshing with the exception of the plants used for morphological characterization. The seed yield was expressed in kg.ha-1 and seeds were stored in a cooling chamber at a controlled temperature of 17 °C and 60 % relative humidity.
The analyses for yield components and plant morphological characterization were done through samples of eight plants per experimental unit, with results expressed as mean values per treatment for each trait. The following traits were studied: plant height (Pht) and height of insertion of the first pod (Hifp) with the aid of a millimeter ruler; stem diameter (Sd), determined through readings of a digital caliper; number of void pods (Nvp); number of single-seed pods (Np1s); number of pods with two seeds (Np2s); number of pods with three seeds (Np3s); number of pods with four seeds (Np4s); pods per plant (Ppp); seeds per plant (Spp); number of secondary branches per plant (Nsbp); seed weight per plant (Swpp) and 1000-seed weight (1000Sw), obtained from eight 100-seed samples as recommended by the Brazilian Rules for Seed Testing-RAS (Ministério da Agricultura, Pecuária e Abastecimiento, 2009).
All seed quality determinations were performed at the Seed Testing Laboratory (STL) at the Federal University of Pelotas (UFPel) and commenced after one month of storage at the STL facility. A germination test (G) was conducted with 200 seeds for each treatment divided into four subsamples of 50 seeds, laid on paper substrate pre-moistened with distilled water in the ratio 2.5 times the mass of dry paper, and rolled to form cylinders that were put in the germination chamber at 25 °C. The temperature, germinated seed counting time, and substrate moisture standards were established according to Brazilian Rules for Seed Testing (Ministério da Agricultura, Pecuária e Abastecimiento, 2009), and the results were expressed as the percentage of normal seedlings. First germination count (FGC) was conducted five days after sowing. Accelerated aging (AE) test was conducted in gerbox-type boxes containing 40 mL of distilled water. Each box had suspended a metal screen on which the seeds were evenly distributed in a single layer. Immediately after laying the seeds on the metal screen the boxes were capped and placed in a BOD incubator at 41 °C, where they remained for 96 hours. After this period, each sample was subjected to the standard germination test, and the first count for normal seedlings performed on the fifth day. Field Emergence (FE) test was performed on small plots seeded to 50 seeds per replicate, performing a single count for a percentage of normal seedlings at day 21th.
Analyses of variance were performed on all data, and when appropriate a polynomial regression was calculated. Data for seed quality recorded as percentage value underwent arc.sen transformation (square root x / 100). Statistical analyses were performed using the Winstat Statistical Analysis System version 1.0.
Results and Discussion
Table 1 shows that results for plant height, stem diameter, number of void pods, and pods with one to four seeds, were not statistically different across population densities. The coefficients of variation for the number of void pods as well as for the number of pods with four seeds were high, which was explained by the individual values recorded, whose small numerical variation between replications resulted, in percentage terms, in major changes.
Table 1: Plant height (Pht), stem diameter (Sd), number of void pods (Nvp), number of single-seed pods (Np1s) and number of pods with four seeds (Np4s) in soybean cultivar BMX Potência RR grown at different plant densities. Means within each column did not differ statistically (p-value < 0.05).
According to Bezerra et al. (2009), the spatial arrangement of plants in the crop is a major factor shaping the competition relationships established among plants, influencing the various morphological attributes, and possibly reflecting on productivity. Yield per plant depends on the arrangement of plant population in the field and the species’ morphological plasticity, the latter dependent on the intrinsic characteristics of the cultivar, such as size, growth habit, plant architecture, and crop management practices.
One of the reasons for the absence of statistical differences on the results of Table 1 may be that the range of plant densities tested fell far below the recommended density. As for the range of values chosen for this work, its justification relied on the need by seed companies for high rates of single-plant multiplication; however, the trends on plant competition on yield components at each plant density showed no impact on the variables presented in Table 1.
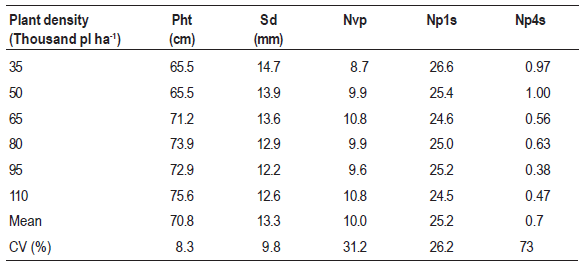
The first pod insertion height varied according to changes in plant density (Figure 1). The insertion of the first pod is an important trait correlated with harvestable yield because it determines the optimum height at which the combine header must be set in order to obtain maximum harvesting efficiency.
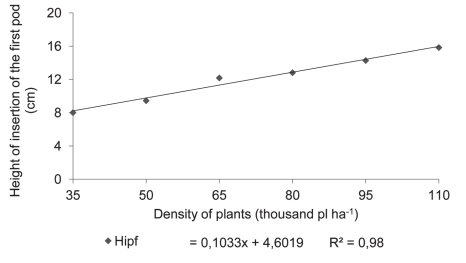
Figure 1: Height of insertion for the first pod (Hifp) in plants of soybean cultivar BMX Potência RR grown at different plant densities.
According to Mauad et al. (2010), the height of the combine header must be adjusted to the height of first pod insertion so that, to minimize harvest losses, this height should be a minimum of 10 cm on a flat surface, and of 15 cm for fields with an irregular surface. In this study, for the 35,000-plant ha-1 population density, the height of insertion of the first pod was of approximately 8 cm.
Increased seeding rates led to an increase in the height of insertion of the first pod for soybean cultivar BMX Potência RR. The two lower densities up to 50,000 plants ha-1 were possibly impaired since they were not able to reach the minimum height to enhance harvesting efficiency. Whenever these plant densities need to be used, adjustments will be necessary to avoid losses. Moreover, pod insertion close to the ground surface could be positively correlated to the total number of pods, impacting on the seed multiplication rate, a trait not so good from the point of view of harvesting efficiency but very attractive for seed production.
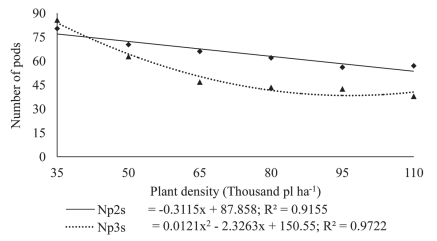
Figure 2: Number of pods with two (Np2s) and three (Np3s) seeds in plants of soybean cultivar BMX Potência RR grown at different plant densities.
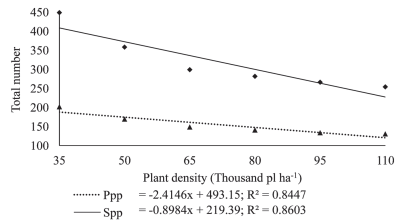
Figure 3: Number of pods per plant (Ppp) and number of seeds per plant (Spp) in plants of soybean cultivar BMX Potência RR grown at different plant densities.
Intraspecific competition for environmental factors such as light, water and soil affect the soybean plant branching capacity. At higher plant densities, there is a lower availability of photo-assimilates for vegetative growth, which will reflect directly on the number of branches (Mauad et al., 2010). This causes the plant prioritize the use of photo-assimilates mostly for the main culm, increasing its length while reducing the emission of lateral branches as shown in Figure 4. Oz (2008), testing four soybean plant densities (280571; 140286; 93523 and 70143 plants ha-1), found that the increase in plant density led to increased plant height as well as that of the first pod, while it decreased the number of secondary branches per plant and the number of pods per plant, which agrees with the results obtained in this study.
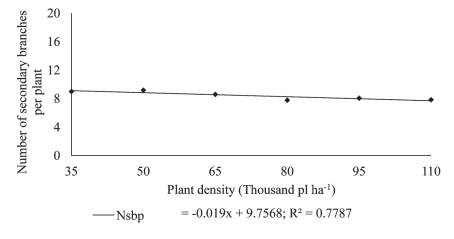
Figure 4: Number of secondary branches per plant (Nsbp) in plants soybean cultivar BMX Potência RR grown at different plant densities.
The number of seeds per plant showed a positive response to decreasing plant densities (Figures 2 and 3). Since the total number of seeds per plant is related to the total number of pods per individual, an increase in the total number of pods will imply a higher number of seeds per plant, a trait also observed by Mauad et al. (2010). This type of response shows that a reduction in plant density increases the multiplication factor, i.e. the production of a larger number of progeny seed per unit seeded.
The reduction in seeding density linearly increased the number of pods per plant (Figure 3). Since at higher plant densities competition for light as a growth factor increases greatly, thus reducing the supply of photo-assimilates, the plant will react to a strategy of producing a smaller number of nodes that in turn will decrease the number of potential branches (Figure 4).
Since nodes are the sites where reproductive buds develop, any reduction in the number of branches implies a lower number of potential nodes, and consequently, a lower number of pods (Mauad et al., 2010). At the lower plant densities, the highest number of pods per plant is explained by the increase in the number of branches (Figure 4), which affects the potential number of nodes and, therefore, a larger number of pods per plant (Figure 3). According to Peixoto et al. (2000), the number of pods is one of the main yield components that is extremely sensitive to variations in plant density. These results agree with those reported by Ludwig et al. (2007), who observed in soybean that the number of pods per plant decreased with the increase in seeding density. Kuss et al. (2008), working with soybean plant densities of 250,000 and 400,000 plants ha-1 in a non-irrigation system, found that the largest population significantly increased grain yield per hectare but decreased the number of pods per plant, while the number of grains per pod remained unaffected.
According to the results recorded in this work, being the multiplication factor (MF) the number of seeds generated by an individual plant, the highest MF corresponded to that of 35,000 plants ha-1, at a ratio of 408:1. The standard plant density, 110000 ha-1, yielded a MF of 227:1. For the data recorded in this experiment, for every 15,000-plant reduction, the MF increased by around 13.3 %. Comparing the extreme plant densities from the range studied in this work, the MF at the lower density was 79.7 % higher than the multiplication factor recorded for the highest plant density.
Seed mass per plant changed according to the seeding density (Figure 5). This can be explained by the fact that at the lower seeding rates a higher number of pods per plant, as well as that of seeds per pod, were observed (Figure 3). Again, under a high plant density growth environment, in which some factors could be in a limited supply, the number of pods per plant and the number of seeds per pod would likely be reduced (Figure 3). Increases in grain yield per plant in response to decreasing seeding rates were also observed by Peixoto et al. (2000), Tourino, Rezende & Salvador (2002) and Procopio et al. (2013).
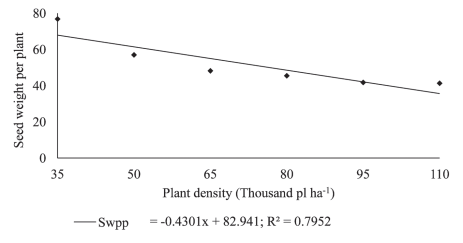
Figure 5: Seed weight per plant (Swpp) in plants of soybean cultivar BMX Potência RR grown at different plant densities.
As shown by these results, soybean plants have the ability to compensate for yield components when grown at lower densities. When analyzing the seed mass per area (Figure 6), it can be seen that at higher densities, even producing a smaller number of pods, seeds or seed weight per plant, the mass gain per area is higher at densities closer to the ideal. In practical terms, for each 1000-plant-per-hectare increase for soybean cultivar BMX Potência RR, there is an increase in seed mass of almost 27 kilograms per hectare up to the highest tested population density.
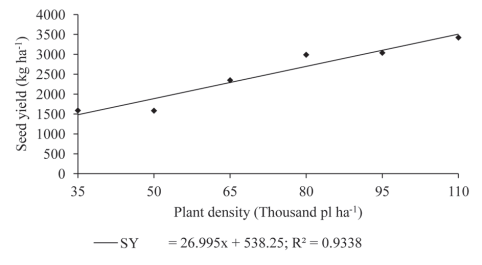
Figure 6: Seed yield (SY) in plants of soybean cultivar BMX Potência RR grown at different plant densities.
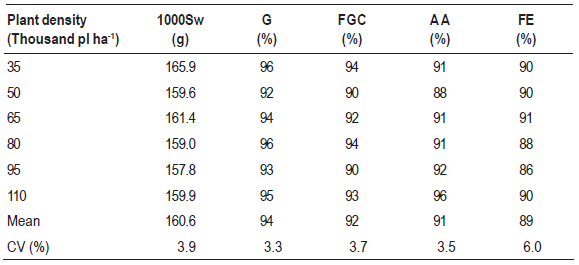
For the variables shown in Table 2, population density had no effect whatsoever, which shows that the physiological quality of seeds does not change at different seeding rates, thus allowing the use of low seeding rates to increase the seed multiplication factor. Vazquez, Carvalho & Borba (2008) also observed no changes on the physiological quality of soybean seeds originated from low plant-density crops.
Table 2: Seed quality assessed through thousand seed weight (1000Sw - g), germination value (G - %), first germination count (FGC - %), accelerated aging (AA - %) and field emergence (FE - %) for seeds of soybean cultivar BMX Potência RR grown at different plant densities. Means within each column did not differ statistically (p-value < 0.05)
The absence of negative effects on seed quality of low plant density crops allows for the use of this strategy as an acceptable option when the objective is to maximize the number of seeds per plant (MF), however, not when the focus is to optimize seed yield per unit area. According to Heiffig et al. (2006), soybean tolerates wide variations in plant population by resorting to morphological changes rather than to variations in grain yield. Therefore, the use of low seeding densities is an important tool that must be restricted to plant material of high economic value, which is the case for promising materials from plant breeding programs.

