











 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo

 Permalink
Permalink
1. Introduction
The genus Conyza Less. belongs to the Asteraceae or Compositae family, it consists of approximately 100 species and its origin is the American continent1. It includes annual/biennial and perennial species that are distributed in temperate and subtropical areas2)(3. They have ruderal habits, of primary succession4 and usually grow in vacant lots, roadsides and railroads. They have also been cited as weeds in pastures and perennial crops, adapting to these little disturbed environments since they are sensitive to soil removal5. These species were introduced on several continents with a wide distribution in Europe, Australia, Africa and Asia3.
The Conyza species are herbs with simple stems or branched from the base, alternate and sessile leaves, of variable margin, from whole to pinnatisect. The generally pedunculated capitulums are arranged on their own or in secondary inflorescences, they have compressed, glabrous or laxly sericeous achenes with papus formed by a series of sub-equal scabrous bristles6)(7.
Twenty-three species of Conyza have been identified in Argentina, which are commonly known as rama negra, mata negra or carnicera, among others1)(8. In the province of Buenos Aires, 9 species were cited: C. blakei (Cabrera) Cabrera; C. bonariensis (L.) Cronquist; C. laevigata (Rich.) Pruski; C. lorentzii Griseb.; C. monorchis (Griseb.) Cabrera; C. pampeana (Parodi) Cabrera; C. primulifolia (Lam.) Cuatrec. & Lourteig; C. serrana Cabrera and C. sumatrensis (Retz.)7.
The Conyza species are annual, although they can behave as biennials, depending on the characteristics of the environment. Most seeds germinate during fall, however, the emergence period extends until the end of spring, so the emergence is staggered in this period9. Various studies in Argentina have demonstrated the existence of two emergence periods for these species, early fall and early spring5)(9)(10)(11. Cardinal germination temperatures (base, optimal and maximum temperature) are unknown in most species of the genus Conyza12. In C. albida, C. bonariensis and C. Canadians, germination was registered in a temperature range of 10-25 °C, with the optimum temperature being close to 20 °C in C. bonariensis12)(13. Seedlings that emerge in the fall and vegetate during the winter months at the rosette stage bear fruit towards the end of spring in response to the photoperiod, elongating the flower stem (a state known as bolting)14. In C. bonariensis and C. canadensis, the stem elongation and the beginning of flowering respond to the photoperiod, taking place in fewer days under long-day conditions13. Flowering induction can take 14 to 22 weeks, depending on the time of germination, so plants that emerge in autumn require more days than those of spring emergence15.
Species of the genus Conyza are considered highly invasive weeds due to their ability to colonize different environments16. Although there are several species in the afore mentioned genus, not all of them behave like weeds in crops, nor do they cause the same loss level5)(17. At world level, they are considered as weeds in more than 40 crops from 70 countries3)(16)(17. The most outstanding species as harmful plants worldwide, both in annual and perennial crops, are Conyza bonariensis and C. canadensis (L.) Cronquist18. The first is native to South America and is abundantly present in Argentina, Brazil, Paraguay, and Uruguay. It is also found in Colombia and Venezuela19. While Conyza canadensis is native to North America and is present on all continents except the Antarctic18.
They have been cited as weeds in grazing systems, as well as in extensive crops, including corn, soybeans, sorghum, cotton, chickpea, wheat and alfalfa20. They were also reported infesting perennial crops, adapting very well to these little disturbed environments5)(21. Their presence is also important in fallows, since they use the water stored in the soil during this period, causing more damage to the crop22. On the other hand, in Argentina before the eighties, the “black branch” species were only problematic in pastures, since conventional tillage was used in areas with intensive and extensive cultivation not allowing them to prosper23.
Certain characteristics such as high fecundity, small seed size, and anemophilous dispersion, among others, have favored its implantation in minimum tillage systems24. Thus, they become very important as weeds in fallow and annual crops under Direct Sowing systems (ds)3. In Argentina, around 90% of the cultivated soybean surface is carried out under ds, and in the last campaigns, the “hairy fleabane” species were the most important weeds of the crop23. The presence of weeds in the crop causes yield decreases mainly due to competition for water, light, and nutrients. Also, they can cause a decrease in grain quality and an increase in the costs of harvest25.
Bedmar and others26 observed losses that oscillated between 20% of the yield with densities of 5-10 plants/m2, and 90% with densities greater than 60 plants/m2 in soybean crops in the southeast of Buenos Aires with the presence of Conyza. Likewise, internationally, yield decreases that range between 22 and 83% have been reported in soybean crops, depending on the level of infestation25)(27.
Herbicide modes of action used for Conyza control include glycins (glyphosate), hormonal (2,4-D> fluroxipir> dicamba> picloram), triazines (atrazine, promethrin, and metribuzin), inhibitors of als enzyme (metsulfuron, chlorimuron), among others, which in many cases are used in mixtures to improve their action11)(12)(24. In addition, there are effective non-chemical methods for controlling species of the genus Conyza, such as tillage and the use of grass coverings5)(28.
The sensitivity of “black branch” to different herbicide modes of action is conditioned by the plant size, dose, time of application and species. Therefore, the opportunity for treatment is key in the efficient and effective management of this weed29. Chemical control is most effective in the early rosette stage20. In other words, as plant height increases, chemical control decreases, whatever the mode of action used. The inflection point for this fall seems to be 15 cm of height, coinciding with the shift from vegetative to reproductive state (stem elongation)5. One of the possible causes would be the greater accumulation of biomass in roots, which provides the plant with reserve energy for subsequent regrowth after chemical control5.
Several factors, such as the inappropriate use of herbicides, the incorrect identification of species and the lack of monitoring, among others, explain why the species of “black branch” have become a growing problem in production systems under ds. Therefore, knowledge of its biology and the adaptive response to various cultural practices, such as crop rotation or increased management intensity, is essential17)(30. For this reason, the objective was to analyze parameters that affect the reproductive success (germination %, fecundity), and the optimal moment of control (height and determination of phenological stages) of Conyza blakei, C. bonariensis, C. lorentzii and C. sumatrensis.
2. Materials and Methods
The analyzed species were Conyza blakei, C. bonariensis, C. lorentzii and C. sumatrensis. Their identification was conducted using the keys of the Flora of the Province of Buenos Aires6, the Prodrome of the Phanerogamic Flora of Central Argentina31, and the Vascular Flora of the Republic of Argentina7. The seeds were harvested manually during December-January-February 2014/2015 in a ruderal environment within the premises of the Agricultural Experiment Station (AES) inta, Balcarce (37° 45' S; 58° 18' W). The seeds were kept at room temperature (18-20 °C) until sowing, which took place on May 5, 2015. For this, 3-liter black polypropylene pots were used, with a mixture of 4 parts of soil and 1 part of sand as a substrate, covering the surface with 5 cm of sterile soil to prevent germination of other species. They were kept outdoors and watered periodically to keep them in optimal humidity conditions. Thirty-five individuals in pots according to a randomized complete block design (rcbd) were analyzed for each species. In turn, a growth chamber test was carried out to determine the germination percentage in the 4 species. One-hundred seeds were placed in Petri dishes, making 3 repetitions for each one, and kept in a growth chamber at 22-25 °C, and with a photoperiod of 12 h of light32. The number of emerged plants was counted at 7, 14 and 21 days after sowing. Germination was evaluated for 21 days since staggered germination has been reported for these species.
Observations were made on each population every 2/3 weeks to record the beginning of the different phenological stages (emergence-rosette stage-elongation-flowering-senescence), which were established when half of the plants analyzed reached this stage.
At the end of each phenological stage, the accumulated thermal time was calculated (thermal time = average daily temperature - base temperature), using as the base development temperature that quoted for C. bonariensis (4.2 °C)12. Temperature and rainfall records of the meteorological station of the aes inta Balcarce were collected.
The plants' height was measured (from the base to their apex) on three sampling dates (09/12/2015, 12/01/2016, 02/02/2016).
The “number of capitulums/plant” and the “number of achenes/capitulum” were quantified to calculate the fecundity of the species with the following formula:
fecundity = No. of capitulums* (No. of achenes/capitulum)
Characters with a high level of significance were analyzed using univariate statistical programs (anova) and the means were compared with the least significant difference test (lsd) (p<0.05). For data analysis, infostat statistical software33) was used.
3. Results and discussion
3.1 Germination
The germination percentage presented significant differences between the evaluated Conyza species, at the different sampling dates. The species with the highest germination percentage at 21 days was C. sumatrensis (55%), in second place C. bonariensis and C. lorentzii with very similar behavior (44 and 43.5% respectively). Lastly, C. blakei was the one with the lowest germination percentage (31.6%). In general, the number of emerged plants increased with the number of days (Figure 1). The determination of the germination percentage will provide vital information to explain the invasion of these species in different environments. Thus, complementary studies of the viability and dynamics of seed emergence together with the competitive ability of the species could explain the differences registered on presence and abundance of the different species in different environments34.
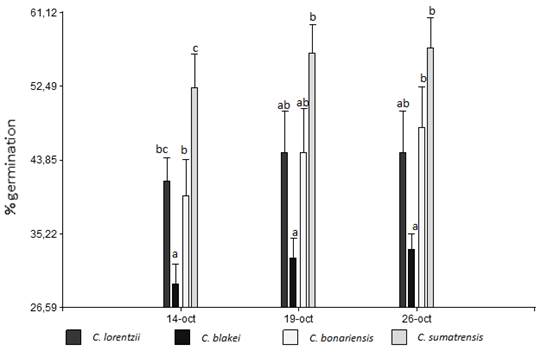
Figure 1: Germination percentage of Conyza lorentzii, C. blakei, C. bonariensis and C. sumatrensis under controlled conditions (for each sample date equal letters indicate non-significant differences according to the lsd test (α= 0.05)). Vertical bars represent the standard error of the mean.
Nandula and others35 obtained 61% germination in the species C. canadensis with a thermal alternation of 24/20 °C and 13 hours of light. Meanwhile, Yamashita and Guimaraes36 reported 90-93% germination in C. canadensis and 78-82% in C. bonariensis with a temperature of 25 °C and a photoperiod of 12 hours of light. Hao and others37 studied the germination percentage under different pollination treatments in the species C. sumatrensis (cross-pollination, self-pollination in the same section and self-pollination with the same individual), resulting in more than 90% germination, in all cases. The values cited in the bibliography are higher than those obtained for C. bonariensis and C. sumatrensis in the present study, while there are no references for the other studied species. The seeds used in the experiment were collected in the field, so they could have suffered stress that limits their germination and/or present some type of dormancy. However, it would provide information about the behavior of the species in coincidence with the first germination flow registered in the field in the evaluated area.
The emergence of the species of Conyza in the field was studied in the southeast of Buenos Aires, establishing 70% of emergence in the months of April and May. During the spring, a second emergence peak occurred, not exceeding 20% of the total emergence9.
3.2 Height
The height of the four Conyza species showed significant differences in the different dates evaluated (Figure 2). Conyza bonariensis was the species that began the stem elongation earlier, reaching 65 cm of final height. Meanwhile, C. sumatrensis and C. lorentzii began to elongate in mid-November, reaching, however, the highest final height (80 and 76 cm, respectively). While C. blakei with 59 cm was the species with the lowest final height and the slowest growth. These data do not agree with the final height cited in the bibliography, being C. sumatrensis the species with the highest height at 150-200 cm, followed by C. bonariensis (100 cm), C. blakei (80 cm) and finally C. lorentzii (60 cm)7. Metzler, Puricelli, and Papa5 recorded a maximum height of 120 cm in C. bonariensis, and of 200 cm in C. sumatrensis. The values cited are higher than those recorded in the present study, where the pot capacity could have limited this variable.
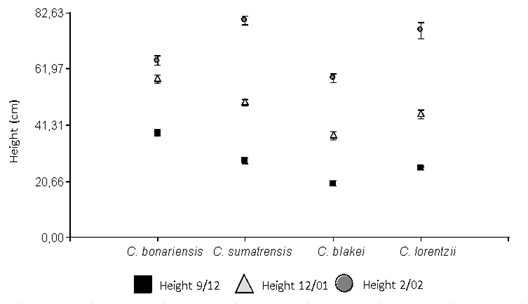
Figure 2: Height (cm) of Conyza lorentzii, C. blakei, C. bonariensis and C. sumatrensis on different sampling dates. Equal letters indicate non-significant differences according to the lsd test (α= 0.05). Vertical bars represent the standard error of the mean.
The Conyza height is very important when establishing management guidelines, since their sensitivity to herbicides is closely linked to this character, in accordance with the transition from vegetative to the reproductive stage and the stem elongation. It has been proved in various tests that once the rosette stage has passed, the plant becomes more tolerant to chemical treatments; this coincides with a height of 15 cm or more, when herbicide controls become difficult, losing efficacy23.
Consequently, C. bonariensis would be the species that requires earlier control, since at the end of October (TT = 1300 °C) it was reaching 15 cm in height. Species C. sumatrensis and C. lorentzii showed an intermediate response, reaching that height on November 7-10 (TT = 1400/1446 °C respectively), while C. blakei was the latest (December 5-10, TT = 1800 °C). The species evaluated in the present study colonize different environments, thus, C. sumatrensis is mainly present in soybean crops, fallows, and ruderal environments, while C. bonariensis and C. blakei invade pastures, being C. lorentzii a more ruderal species34. The thermal time accumulated in each stage is relevant to design management strategies by environment, according to the predominant species. In this way, for example, if control is required in a pasture environment, it should be carried out earlier compared to that required in a soybean crop, since in the first one it is C. bonariensis the predominant species, while in soybean C. sumatrensis was registered as the predominant species34.
3.3 Reproductive Character Analysis
The first species to flower was C. bonariensis, reaching this state on November 20 (27 weeks after germination, TT = 1586 °C) (Figure 3). Secondly, the flowering of C. blakei was recorded on January 27 (37 weeks after germination, TT = 2747 °C). Finally, during the month of February, the species C. sumatrensis and C. lorentzii started flowering on February 10 and 18, respectively (39 and 40 weeks after germination, TT = 2997/3151 °C respectively). Zinzolker and others13 analyzed the phenology in C. bonariensis and C. canadensis, recording that stem elongation and the beginning of flowering occurred in fewer days under long-day conditions. It was recorded that in optimal thermo-period conditions (27-22 °C) C. bonariensis takes 11 weeks to lengthen the stem and an additional 3 weeks to flower16. In this case, the accumulated thermal time is around 1960 °C ((24.2-4.2) * 14 weeks * 7 days), similar to that recorded in this study. During the 2014-2015 campaign, the flowering date of the four species of Conyza analyzed in the present study34 was recorded in the town of Balcarce (average annual temperature 13.3 °C). In this study, the earliest flowering species was C. bonariensis (December 10-15), which also had a longer flowering period, extending until April. Conyza sumatrensis and C. blakei began flowering two months after the previous one (February 10-15), while C. lorentzii was the last recorded species, February 20. Although the dates were similar for C. sumatrensis and C. lorentzii, they differed for C. bonariensis and C. blakei, being earlier in the year 2015-2016 and under the study conditions. Metzler, Puricelli, and Papa5 recorded in Parana (average annual temperature 18.2 ºC), Entre Rios, the flowering of C. bonariensis in mid-October and C. sumatrensis at the end of January. The later flowering in southeast Buenos Aires could be due to the lower temperature compared to the province of Entre Rios.
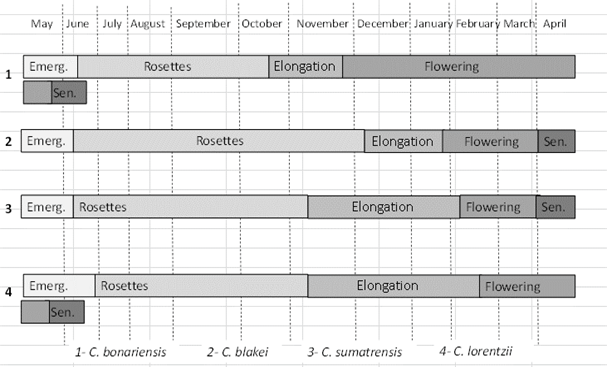
Figure 3: Life cycle of Conyza bonariensis, C. blakei, C. sumatrensis and C. lorentzii in southeastern Buenos Aires (2015-2016).
The senescence of the 4 species occurred within the first year of growth, behaving like annual species. The senescence of C. sumatrensis and C. blakei occurred during the first week of April 2016 (TT = 3800 °C), being the last week of May of the same year for C. bonariensis and C. lorentzii (TT = 4250 °C). Although all species are referred to as annual in the bibliography, it is known that depending on the time of emergence they can behave as biennials when they emerge during spring17)(21)(23.
In addition, the accumulated thermal time was calculated for the 4 species of Conyza at the end of each phenological stage (Table 1). Such data will be very useful to consider the duration of each stage in different geographical areas.

3.4 Number of capitulums per plant
The number of capitulums per plant presented significant differences in the different species of Conyza analyzed. The highest production of capitulums was observed in C. lorentzii and C. blakei, with an average number of 1,202 and 1,056 capitulums per plant, respectively. C. sumatrensis and C. bonariensis showed significant differences (α = 0.05) with the previous group with 529 and 415 chapters, respectively (Figure 4).
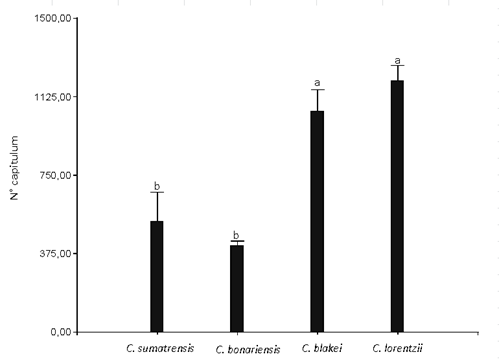
Figure 4: No. of capitulums of Conyza bonariensis, C. sumatrensis, C. blakei and C. lorentzii (equal letters indicate non-significant differences according to the lsd test (α= 0.05)). Vertical bars represent the standard error of the mean.
The maximum number of capitulums per plant found in C. bonariensis matches the 400 capitulums/plant recorded by Wu and others12 under laboratory conditions. Yet, it is higher than those recorded by Kempen and Graf38, and Green and others20, who reported an average of 290 and 232 capitulums/plant, respectively. Zambrano Navea21 registered the number of capitulums in C. bonariensis for different cohorts, born in the field during fall, recording a decrease in the number of sections with the emergence delay (200-83-30 in 2010 and 184-58-38 in 2011-2012), although the number of achenes per section did not show significant differences in the different groups.
3.5 Number achenes/capitulum
The number of achenes per capitulum showed significant differences in the different evaluated species (Figure 5). The behavior of this variable was inverse to that observed in the production of capitulums/plant, being C. bonariensis (276.72 achenes/capitulums) the species that produced the highest number of achenes, second C. sumatrensis (138.3 achenes/capitulum), and finally, C. blakei (66.4 achenes/capitulum) and C. lorentzii (53.04 achenes/capitulum), not showing a significant difference between them.
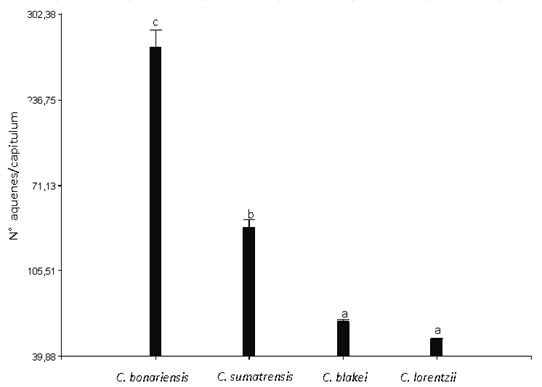
Figure 5: No. of achenes/capitulum of Conyza bonariensis, C. sumatrensis, C. blakei and C. lorentzii (equal letters indicate non-significant differences according to the lsd test (α= 0.05)). Vertical bars represent the standard error of the mean.
Diez de Ulzurrun and others34) determined the number of achenes/capitulum in the same species, under natural conditions, during the 2014-2015 campaign. The species with the highest prolificacy was C. bonariensis (178.5 achenes/capitulum), followed by C. sumatrensis (105.7 achenes /capitulum), C. blakei (73.8 achenes/capitulum) and C. lorentzii (46.9 achenes/capitulum). Although they are slightly lower than those recorded in this study, the decreasing order of the species according to the production of achenes is preserved. Similarly, Urdampilleta and others1 recorded an average of 83.4 achenes/capitulum in C. blakei, 226.4 achenes/capitulum in C. bonariensis and 142 achenes/capitulum in C. sumatrensis.
3.6 Fecundity
This variable showed significant differences between the 4 Conyza species analyzed, being C. bonariensis the species that showed the highest fecundity, with an average of 114,838.8 achenes/plant, followed in order of importance by C. sumatrensis (73,176.5 achenes/plant), C. blakei (70,139.5 achenes/plant) and C. lorentzii (63,785.9 achenes/plant), these last did not differ significantly between each other (Figure 6). The species with the longest cycle duration were C. bonariensis and C. lorentzii (TT = 4200 °C), however, this longer duration and, consequently, resource uptake was not associated with higher fecundity in the second species.
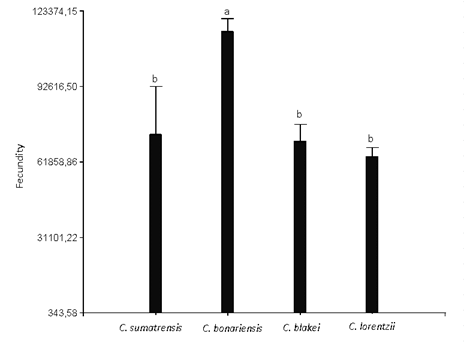
Figure 6: Fecundity of Conyza sumatrensis, C. bonariensis, C. blakei and C. lorentzii (equal letters indicate non-significant differences according to the lsd test (α= 0.05)). Vertical bars represent the standard error of the mean.
In Spain, Zambrano Navea21 recorded early cohorts of C. bonariensis with an average fecundity of 86,066 achenes/plant, which is significantly lower than that recorded in the present study. However, they are similar to those obtained by Wu and others12) under controlled irrigation and temperature conditions, who recorded 119,100 achenes/ plant. The C. sumatrensis species in field conditions had an average fecundity of 60,000 achenes/plant, being slightly lower than those registered in the present study37. Moreover, Metzler, Puricelli and Papa5 estimated in this species a production of 30,000 achenes/plant at low densities. However, Gianelli and others14 estimated a production of 3,418,293 achenes/plant with a density of 1 plant/m2 in C. sumatrensis, confirming a greater distribution of biomass in terms of allocation to reproduction (biomass of reproductive organs/total biomass) at low densities.
The data recorded in this study, while preliminary, will be of interest in planning environmental management strategies, taking into account the distribution of these species in the southeast. It could also be validated for other zones using the tt required for each species. Furthermore, it will provide valuable information for species C. blakei and C. lorentzii, limited in the national/international literature.
4. Conclusions
The germination percentage presented significant differences between species, being Conyza sumatrensis the species with the highest number of births.
The different phenological stages were established for the four species of Conyza, being C. bonariensis the species with the earliest elongation, the first to reach the reproductive stage and the one with the highest fecundity.
All species completed the life cycle within one year from germination.
Under the study conditions, the species with the greatest reproductive success were C. bonariensis and C. sumatrensis, the first requiring earlier controls.

